





There is now a consensus, based on histological, biochemical and spectral absorption data, that the yellow colour observed at the macula lutea is a consequence of the selective accumulation of dietary xanthophylls in the central retina of the living eye. Scientific research continues to explore the function(s) of MP in the human retina, with two main hypotheses premised on its putative capacity to (1) protect the retina from (photo)-oxidative damage by means of its optical filtration and/or antioxidant properties, the so-called protective hypothesis and (2) influence the quality of visual performance by means of selective short wavelength light absorption prior to photoreceptor light capture, thereby attenuating the effects of chromatic aberration and light scatter, the so-called acuity and visibility hypotheses. The current epidemic of age-related macular degeneration has directed researchers to investigate the protective hypothesis of MP, while there has been a conspicuous lack of work designed to investigate the role of MP in visual performance. The aim of this review is to present and critically appraise the current literature germane to the contribution of MP, if any, to visual performance and experience.
En la actualidad, en función de los datos histológicos, bioquímicos y de la absorción espectral, se ha alcanzado un consenso, de que el color amarillo observado en la mácula lútea es consecuencia de la acumulación selectiva de xantófilos dietéticos en la retina central del ojo vivo. La investigación científica continúa examinando las funciones del pigmento macular en la retina humana, con dos hipótesis principales formuladas sobre su supuesta capacidad para: 1) proteger la retina frente a la lesión (foto)oxidativa por medio de sus propiedades de filtración óptica y/o antioxidantes, la llamada hipótesis protectora e 2) influir en la calidad del rendimiento visual por medio de la absorción selectiva de luz de longitud de onda corta antes de su captura por parte de los fotorreceptores, lo que atenúa los efectos de la aberración cromática y dispersión de la luz, la llamada hipótesis de la agudeza y la visibilidad. La epidemia actual de degeneración macular relacionada con la edad ha dirigido a los investigadores a examinar la hipótesis protectora del pigmento macular, mientras que es evidente la falta de investigación destinada a investigar el papel del pigmento en el rendimiento visual. El objetivo de la presente revisión es describir y valorar de forma crítica los estudios publicados actuales pertinentes a la contribución del pigmento macular, si desempeña algún papel, en el rendimiento y experiencia visual.
Vision, and how we perceive the world, involves the complex interaction of physical, physiological and psychological processes, which ultimately provide the final sensation of seeing. “Visual performance”, as discussed here, describes the sensitivity of the eye where limits of vision are quantified using established clinical and laboratory techniques. Such techniques cannot readily account for variable and highly individual experiences and interactions in the real world. “Visual experience” incorporates subjective experience, which may, for example, explain inconsistencies between patient's symptoms and measured functional vision. Any influence of macular pigment (MP) on vision needs to be assessed therefore, in terms of both measured performance and reported experience.
Macular pigment was first observed by Buzzi 1 in 1782, and speculation persists as to its role in the visual system. Indeed, at first there were conflicting views as to the very existence of this pigment in the living eye, with numerous authors, including Home 2 and Gullstrand 3 believing it to be a post-mortem artifact.
It has long been recognised that MP preferentially absorbs short wavelength light prior to photoreceptor stimulation, and the hypothesis that filtering such defocused short wavelength light could enhance visual performance by reducing the effects of chromatic aberration goes back as far as Schültze 4 in 1866. This hypothesis, especially in relation to MP, remains unproven and poorly investigated. In this review, we explore the contribution of MP to visual performance and experience, and report and critically appraise the evidence in support of the notion that MP is important for vision.
The selective accumulation at the macula of only three dietary carotenoids, to the exclusion of the other forty dietary carotenoids, suggests an exquisite biological selectivity for lutein (L), zeaxanthin (Z) and meso-zeaxanthin (meso-Z) at the site of maximum visual acuity in the human retina, and also suggests a specific role for these carotenoids which is uniquely suited to this anatomic location. Given that Darwinian natural selection is based on the premise that phenotypic expression of genetic background confers advantage before and until the period of procreation, it is reasonable to infer that the biological selectivity of MP's accumulation in the retina is advantageous in young and middle age.
MP may protect against the development of age-related macular degeneration (AMD) by defending the retina against cumulative and chronic (photo)-oxidative damage. It is likely, however, that the primary role of MP rests on its contribution to visual performance and experience, although the pigment may also longitudinally contribute to the preservation of macular function by preventing or delaying the onset of retinal disease such as AMD through its protection against chronic (photo)-oxidative damage. In other words, and in theory at least, MP's putative contribution to visual performance rests on its optical properties, whereas the putative protective effect of this pigment for AMD rests on its optical and/or its biochemical properties.
MP alters the spectral composition of the light incident upon macular photoreceptors, but whether such short wavelength absorption influences the quality of the visual experience, and whether the magnitude of any such effect correlates with MP optical density (MPOD), are questions that remain unanswered. Meaningful comment on the contribution of MP, if any, to visual performance must (1) consider the primary factors that affect visual performance, (2) outline the properties of MP that make it potentially important for visual performance in light of any such limiting factors, (3) critically appraise the current literature germane to the role of MP in visual performance and experience and (4) suggest experimental strategies designed to investigate whether MP is important for visual performance and experience.
Visual performanceCurrent and unifying conceptsSnellen was the first to standardise the measurement of visual acuity with his letter chart, a chart design, which despite numerous limitations, remains the most widely used means of quantifying visual performance in the clinical setting. There remains, however, a myriad of other independent and/or overlapping techniques by which one can measure visual performance and experience across a range of functional levels.
Vision includes the capacity to detect objects against a contrasting background, to detect gaps between objects, to perceive subtle vernier offsets (which provides one example of hyperacuity), to recognise and identify objects, to perceive colour, to detect movement, and to perceive depth, amongst other faculties. It is important to note that the capacity to recognise a small distant object bears little relation to the capacity to differentiate colours, or to detect a potential threat such as an oncoming vehicle in the peripheral field of view.
Visual performance is critically dependent on illumination, and the range of illumination we experience in the course of a typical day is vast. The visual system copes with such changes in illumination by adapting to the prevailing conditions, and can function through an approximate 8 log unit luminance range. Although adaptation facilitates performance over a wide range of ambient illumination levels, it does not follow that we see equally well at all levels. Under dim conditions, for example, the visual system is very sensitive and can detect subtle changes in luminance, but acuity for pattern details and colour discrimination is poor.
Shlaer5 has explored the relationship between illumination and visual acuity (Figure 1). Converting his findings to Snellen equivalent, daylight (photopic) performance of 20/10 reduces to 20/600 under dim conditions, a 60-fold reduction. Threshold visibility, colour appearance and visual acuity all vary dramatically with illumination, and these visual parameters change over the time-course of light and dark adaptation. Therefore, and by definition, no single test or testing condition can be used to investigate visual performance, and no single test can predict performance on other tests.
 and retinal illuminance. As retinal illuminance increases, visual acuity increases by up to 2 log units (cone-mediated improvements account for the most significant improvements from approximately 6/60 to 6/3 Snellen equivalent-see upper portion of curve). Shlaer5.")
Relationship between visual performance (as log visual acuity) and retinal illuminance. As retinal illuminance increases, visual acuity increases by up to 2 log units (cone-mediated improvements account for the most significant improvements from approximately 6/60 to 6/3 Snellen equivalent-see upper portion of curve). Shlaer5.
Further, any discussion of the visual processes must include those mechanisms contributing to perception. The visual system employs numerous anatomic and physiological strategies, including lateral interactions between cells, specific receptive field organisation, spatial retinotopic organisation in retinal and non-retinal areas of the pathway, colour opponency and parallel visual pathways, amongst others, in order to achieve an instantaneous, coherent and highly detailed perception of the outside world and our position within it. Such image processing is not exclusive to the brain, but extends throughout the visual pathway beginning at the retina.
The eyes and brain are thus inextricably linked with the visual universe. The eyes actively record the form, colour and movements of the world, and the brain moulds these raw perceptions into recognisable patterns. The retina essentially acts as a spatial, temporal and spectral filter of patterns of light striking its surface. Its anatomic structure and the functional properties of individual cells determine the type of information extracted from a visual scene and delivered to the brain.
Specialisation of the maculasThe macula, which comprises less than 4 % of the total retinal area, subserves almost all of our useful photopic vision. Several distinctive anatomic and neural adaptations facilitate such a high level of visual performance. These include:
- 1.
Cone density peaks at the centre of the macula (fovea), which intersects the line of sight. Cones here are smaller, more densely packed and more numerous than elsewhere in the retina, thus extending the limits of spatial acuity. Cone density exceeds rod density only at the lower part of the foveal slope, reaching a maximum at the base of the fovea (foveola) where cone density is over three times that observed at the foot of the foveal slope. 6 Rods, ganglion cells and all inner nuclear layer neurons are absent from the foveola, so that only here is light directly incident on photoreceptors (elsewhere light must traverse the various retinal cells and layers to reach photoreceptors). It is also worth noting that short wavelength sensitive cones are absent at the foveola.
- 2.
Midget pathways arising from these foveal cones dominate. Such parvocellular midget pathways are tuned to high spatial frequencies and also exhibit colour opponency.
- 3.
Such midget pathways are distinctive because of the absence of convergence of photoreceptor signals onto bipolar and ganglion cells. Absent or reduced convergence of information preserves the data gathered at the fovea for delivery to the visual cortex. Such differences between foveal and extra-foveal pathways generate a hierarchy in the processing of information gathered by the retina.
Anatomic and physiological observations, such as the differential light sensitivity of photoreceptors, the variable density and distribution of photoreceptors and ganglion cells across the retina and the convergence of information from the extra-foveal retina, means that a hierarchy exists in the architecture of retinal processing, where foveal information is given higher priority. This hierarchy is preserved to the striate cortex, where a high percentage of cortical cells are dedicated to information of foveal origin. The central retinal pathways have by far the greatest proportion of representation (estimates range from 25 % of the cortex devoted to the central 5 degrees, 37 % devoted to the central 15 degrees 7 and 87 % of the cortex devoted to the central 30 degrees of visual field 8).
Having outlined those anatomic and neural factors central to primates’ capacity for high acuity vision, it is now important to consider the potential role of MP in visual performance. In order to do so, it is essential to characterise (a) the optical limitations that might restrict visual performance (in particular chromatic aberration and light scatter) and (b) the properties of MP that might serve to lessen the effect of such limitations, and thereby facilitate optimal visual performance.
Optical limitations of the eyeMonochromatic aberrations and diffraction limit the image quality produced by the eye, so that the image is not always a high quality representation of the object. While there is significant ocular and neural correction for, and adaptation to, such image defects, MP most likely has no role in altering their effects (although Kvansakul et al. 9 have noted some surprising observations of a trend towards lower root mean square wavefront aberrations in a small group of subjects following supplementation with L and Z, which, they postulate, may be as a result of the as yet unknown effects of carotenoid intake on crystalline lens function).
Chromatic aberrationChromatic aberration, comprising both longitudinal (LCA) and transverse (TCA) components, has been cited as possibly the most significant aberration affecting visual quality. 10 Indeed, LCA creates up to two dioptres of wavelengthdependent optical defocus. Campbell and Gubbisch 11 have demonstrated improvements in contrast thresholds of up to 65 % at intermediate spatial frequencies once monochromatic yellow light is employed in place of spectrally broadband white light. Although Bradley 12 later modelled the effects of chromatic aberration, and concluded that the effect of chromatic aberration on the modulation transfer function was small, and equivalent to approximately 0.15D of defocus, upper resolution limits of the visual system however, are most likely defined by the effects of chromatic aberration. 13
The effect of LCA across wavelength, in terms of blur, is non-linear, as shorter wavelengths are significantly more defocused than longer wavelengths. For example, an eye focussed at 550nm, light at 460nm suffers 1.2D myopic defocus, while the equivalent long wavelength of 640nm is only 0.50D out of focus. 10 This serves to create a purple blur circle haze around the focussed “green” component. Figure 2 demonstrates the non-linearity of defocus and the relative luminance profile across wavelength. As the spectral extremities have less luminosity, the effects of chromatic aberration on image focus are mitigated in terms of the effects on vision. Mitigation is potentially further aided by the fact that blue light is selectively absorbed by MP.

Illustration of the relative luminance profile and the effect of chromatic aberra tion across wavelengths. The relative blur is more pronounced at the blue end of the spectrum such that, for example, the short wave 460nm text is significantly more difficult to recognise than the long wave 640nm text for the above scenario where the optimal focus is between 540-560nm.
If one looks up to the sky on a bright, cloudless sunny day, one could be fooled into thinking that the sun's rays traverse an unobstructed path to the eye. Furthermore, one could certainly not imagine that the quality of the light visible was being degraded as it traversed the seemingly clear sky, even in the most remote countryside locations far from the smog-filled cityscapes, on its way to the eye. The fact that the sky is blue is testament to the impact of the process of light scatter, whereby particle matter abstracts and re-radiates energy from light incident upon it.
A multitude of visible and non-visible particles, varying in size from atmospheric oxygen and nitrogen, to haze aerosols, to larger complexes such as fog, cloud and rain, all contribute to such scatter. Wooten & Hammond, 14 in an excellent review of the importance of light scatter to the “visibility” of objects, eloquently describe why light scatter, especially that induced by haze aerosols “critically determines how far one can see and how well details can be resolved”, so that, aside from the optical and neural limits, “scatter in the aerosol haze is the primary determinant of visual discrimination and range in the outdoors”.
The question therefore arises, what effect does light scatter have on visual performance? And it is a good question. On a clear day one can see for miles despite the effects of scatter. Wooten & Hammond, 14 however, propose a model whereby compensation for the effects of light scatter, such as could reasonably be achieved by increasing the optical density of MP, would increase the visibility and discriminability of targets in natural settings. In their model, a 1 log unit increase in MPOD attenuates the veiling luminance of the short-wave dominant background by 26 % (or 17 % for a more practical 0.5 log unit increase in MPOD), while having minimal effect on the short wave deficient distant target. The attenuation of the effects of light scatter is thereby observed to enhance target detection and discrimination capacity, and extend the visual range by up to 18.6 %.
Tackling the question from another perspective, the problems caused by scatter, while not consciously experienced by most people, do become a significant symptom of which many patients complain in the form of discomfort and disability glare. Aside from patients without detectable ocular abnormality, typical patients with such symptoms include those with cataract, corneal abnormalities, intraocular inflammation, and following laser refractive surgery, amongst others. Therefore, scatter does have an adverse effect on the visual experience of normal subjects and on those with ocular pathology, and any means of alleviating such effects would be of clinical importance.
The possible effect of aberrations such as LCA, and also of short wavelength light scatter, is that capacity limits are somewhat reduced so that the anatomic limits of acuity based on foveal cone diameter (30 seconds arc — equivalent to 6/3) are seldom achieved, even in healthy normal individuals, with the exception of hyperacuity tasks which have different underlying neural bases.
So the question arises, what are the properties of MP that might allow it to improve visual performance in light of the limiting factors outlined above?
Optical and anatomic properties of MPMP's optical and anatomic properties have prompted the “optical” hypothesis of this pigment, which has been discussed in detail by Reading & Weale 15 and later by Nussbaum et al. 16 The optical effect of MP is somewhat evidenced by two entoptic phenomena known to exist which are specific to the macula, namely Maxwell's spot and Haidinger's brushes. 16 The former, first described in 1844, is attributed directly to the deposition of pigments at the macula and results in a dark red spot being visible around the fixation point if a brightly illuminated white surface is viewed alternately through purple and neutral filters. Magnussen et al. 17 have shown that the absence of short-wave-sensitive cones in the human foveola, which normally goes unnoticed unless a subject's field of view is restricted to the foveola, producing the artificial colour vision defect of foveal tritanopia, 18,19 results in a blue scotoma which can be visualised as the negative afterimage of a short-wavelength adapting field on a larger white background. The afterimage has an annular shape with a lighter inner region that corresponds to Maxwell's spot, and a small bright spot in the centre, corresponding to the foveal blue scotoma. The MP distribution measured for the same observers closely corresponded to the lighter annular region of the afterimage.
Haidinger's brushes, first reported in 1844, refers to a propeller-shaped image which is seen most clearly through a rotating filter producing plane-polarised light. It is known that lutein has dichroic properties 20,21 and it has been shown that bovine lutein and zeaxanthin bind to bovine retinal tubulin. 22 It is thus possible that dichroic macular pigments are laid down in a highly organised manner following the radial arrangement of Henle's fibres at the macula, thus explaining the shape and brush-like appearance of the propeller-like images. 23
However, it should be noted that neither of these entoptic phenomena is visible in normal viewing conditions, probably because of adaptatory effects, particularly at the level of the visual cortex. It is uncertain whether the concentration of MP has any significant influence on vision under such conditions.
MP may be important for visual performance and/or experience however by at least one of the following mechanisms (summarised by Walls & Judd 24): MP may enhance visual acuity by reducing chromatic aberration (effects); MP may reduce visual discomfort by attenuation of glare and dazzle; MP may facilitate enhancement of detail and visual contrast by the absorption of “blue haze”. MP has the capacity to achieve the above optical effects because of its optical properties and because of its location within the retina.
The term macula lutea is actually attributable to the presence of the xanthophyll pigments, L, Z, and meso-Z at the central region of the retina, which give rise to the appearance of a yellow spot (macula lutea) when viewed under red-free light (Figure 3). The yellow coloration of MP is such that it selectively absorbs blue-green incident light, with maximum absorption circa 460nm and little or no absorption above 530nm. 25 Given that (1) the peak retinal spectral sensitivity lies at 555nm, (2) the proportion of blue (short wavelength sensitive) cones in the central macula is far lower than that of red (long wavelength sensitive) and green (medium wavelength sensitive) cones and (3) the region of maximal visual performance, the foveola, is essentially devoid of short wavelength sensitive cones, it would appear that the optical properties of MP are such that it attenuates the component of light that is least beneficial, and most deleterious, with respect to visual performance and experience. As Wald 26 summarised, the various adaptive mechanisms in the human eye serve to “withdraw vision from the blue” end of the spectrum.
 and in the inner nuclear layer at the parafoveal site (b).")
Histological section illustrating the spatial profile and pre-receptorial location of MP, the main location of macular pigment was in the layer of the fibres of Henle in the fovea (a) and in the inner nuclear layer at the parafoveal site (b).
Two aspects of MP's location within the retina are also central to the hypothesis that it has a role to play in visual performance. Firstly, although MP is found throughout the retina and other ocular structures, 27 it reaches its greatest concentration at the macula, and remains optically undetectable elsewhere. Secondly, and importantly, MP is located at a pre-receptoral level, so that absorption of short wavelength light occurs prior to stimulation of the underlying photoreceptors, thereby altering the spectral distribution of light incident on such photoreceptors in a favourable way (Figure 3).
Short wavelength light absorption attenuates the more disadvantageous component of LCA. Retinal image quality is thereby improved, and visual performance across the full contrast range is theoretically more refined. As MP absorption overlaps with that of rhodopsin, MP may reduce rod signal effectiveness in the mesopic range, and thus extend the usefulness of cone-mediated vision into the mesopic range. 9 In addition, short wavelength light absorption has the benefit of improving target contrast by selectively reducing the scattered short wavelength light in the background. Reduced LCA and reduced scatter effects, resulting from MP's absorptive characteristics, have the potential to improve visual acuity and target visibility, and perhaps in an interactively additive fashion. 14
The higher energy and retinal irradiance associated with shorter wavelengths (International Commission on Non-Ionizing Radiation Protection, 1997 28) also merits consideration. Bright light, which interferes with the quality of visual perception, is termed glare, of which there are numerous types. In high luminance or high contrast situations, where glare and dazzle are maximal, MP absorption of short wavelength light attenuates the highest energy light component, and reduces retinal irradiance, and therefore may minimise the impact of glare on performance, and increa se the threshold for photophobia under normal viewing conditions. Because of their linear structure, L, Z, and meso-Z also exhibit dichroic properties, 29 which facilitate glare reduction by preferential absorption of polarised light. Glare symptoms remain a common and important clinical entity in optometric and ophthalmological practice, and very troublesome for those who experience it. 30 Furthermore, symptoms of glare remain difficult to quantify and treat. Interestingly, difficulty with glare is often one of the earliest manifestations of AMD.
It should now be clear, because visual performance is a complex subject, which is difficult to quantify, and dependent on numerous independent and overlapping variables, that to investigate the contribution of any one factor (such as MP) presents numerous challenges. It is with this thought in mind that currently available evidence on the impact of MP on visual performance and experience will now be explored.
Evidence that MP plays a role in visual performance and experienceBackgroundThe evidence in relation to a role for MP in visual performance is sparse and is largely associative. To our knowledge, there are no published studies which have satisfactorily investigated the hypothesis that MP influences visual performance and experience. However there are numerous and conflicting reports on the effect of yellow filters on visual performance, 31 but none of these have included measures of MPOD. Failure to do so confounds any reasonable interpretation of short wavelength light absorption effects on visual performance, as variations in MPOD between and within study populations could account for the reported observations.
There are thus two strategies to investigate the impact of MP on visual performance. The first is to quantify performance using a range of functional tests, and to correlate the results with measures of MPOD. Given the other variables involved in vision, the true effect of MP would, in our opinion, prove difficult to isolate with such a paradigm. The alternative and most appropriate means to investigate the effect of MP is to measure baseline visual performance, as above, and to record baseline MPOD, and then repeat functional vision tests during an extended period of supplementation with MP xanthophylls. If MP influences visual performance it must do so either as (1) a filter or (2) through some biological mechanism. With respect to the former (1), any effects on visual performance should follow the known absorbance characteristics of the pigments. Hence, the visual stimuli to be used to investigate the role of MP should have significant amounts of short wave energy, in order to replicate the effects of ecologically valid stimuli (e.g. the sun) which have lots of short wave energy. Biological effects (2) would likely be based on either enhanced protection (healthier retinas and crystalline lenses would lead to better vision, especially in the elderly) or effects throughout the visual system. If MP has a role, and its contribution is related to either its optical density and spectral absorbance characteristics, or to possible biological effects on retinal, crystalline lens and visual system health, then increasing MPOD through supplementation should result in improved performance and experience. The key then is to accurately detect and quantify any such changes through a comprehensive battery of appropriate tests that analyse vision on a number of functional levels, including basic acuity, contrast sensitivity across illumination levels, colour perception, and glare sensitivity, amongst others.
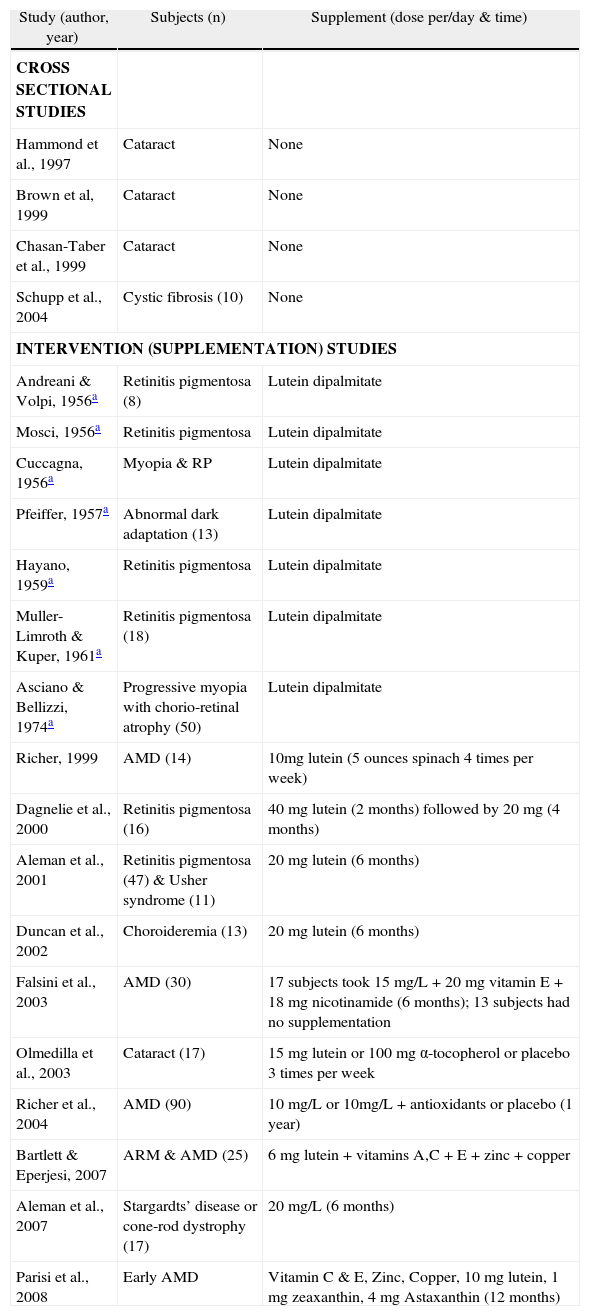
Those studies that have addressed visual performance are largely confined to populations with established eye disease (summarised in Table 1), and therefore the results should be interpreted with full appreciation of the fact that the findings do not necessarily hold true for subjects without retinal pathology. Studies involving normal subjects will therefore be reviewed separately here (summarised in Table 2).
Publications exploring the relationship between macular pigment and visual performance and experience in subjects with ocular disease
| Study (author, year) | Subjects (n) | Supplement (dose per/day & time) |
| CROSS SECTIONAL STUDIES | ||
| Hammond et al., 1997 | Cataract | None |
| Brown et al, 1999 | Cataract | None |
| Chasan-Taber et al., 1999 | Cataract | None |
| Schupp et al., 2004 | Cystic fibrosis (10) | None |
| INTERVENTION (SUPPLEMENTATION) STUDIES | ||
| Andreani & Volpi, 1956a | Retinitis pigmentosa (8) | Lutein dipalmitate |
| Mosci, 1956a | Retinitis pigmentosa | Lutein dipalmitate |
| Cuccagna, 1956a | Myopia & RP | Lutein dipalmitate |
| Pfeiffer, 1957a | Abnormal dark adaptation (13) | Lutein dipalmitate |
| Hayano, 1959a | Retinitis pigmentosa | Lutein dipalmitate |
| Muller-Limroth & Kuper, 1961a | Retinitis pigmentosa (18) | Lutein dipalmitate |
| Asciano & Bellizzi, 1974a | Progressive myopia with chorio-retinal atrophy (50) | Lutein dipalmitate |
| Richer, 1999 | AMD (14) | 10mg lutein (5 ounces spinach 4 times per week) |
| Dagnelie et al., 2000 | Retinitis pigmentosa (16) | 40 mg lutein (2 months) followed by 20 mg (4 months) |
| Aleman et al., 2001 | Retinitis pigmentosa (47) & Usher syndrome (11) | 20 mg lutein (6 months) |
| Duncan et al., 2002 | Choroideremia (13) | 20 mg lutein (6 months) |
| Falsini et al., 2003 | AMD (30) | 17 subjects took 15 mg/L + 20 mg vitamin E + 18 mg nicotinamide (6 months); 13 subjects had no supplementation |
| Olmedilla et al., 2003 | Cataract (17) | 15 mg lutein or 100 mg α-tocopherol or placebo 3 times per week |
| Richer et al., 2004 | AMD (90) | 10 mg/L or 10mg/L + antioxidants or placebo (1 year) |
| Bartlett & Eperjesi, 2007 | ARM & AMD (25) | 6 mg lutein + vitamins A,C + E + zinc + copper |
| Aleman et al., 2007 | Stargardts’ disease or cone-rod dystrophy (17) | 20 mg/L (6 months) |
| Parisi et al., 2008 | Early AMD | Vitamin C & E, Zinc, Copper, 10 mg lutein, 1 mg zeaxanthin, 4 mg Astaxanthin (12 months) |
AMD indicates age-related macular degeneration; ARM, age-related maculopathy; MPOD, macular pigment optical density.
Publications exploring the relationship between macular pigment and visual performance and experience in normal subjects
| Study (author, year) | Subjects (n) | Supplement (dose & time) |
| CROSS SECTIONAL STUDIES | ||
| Hammond et al., 1998 | Normals | None |
| Stringham et al., 2003 | Normals | None |
| Stringham et al., 2003 | Young normals (16) | None |
| Stringham & Hammond, 2007 | Normals (36) | None |
| Engles et al., 2007 | Normals (80) | None |
| INTERVENTION (SUPPLEMENTATION) STUDIES | ||
| Monje, 1948a | Normals (14) | Lutein dipalmitate (2-6 months) |
| Wustenberg, 1951a | Normals (7) | Lutein dipalmitate |
| Klaes & Riegel, 1951a | Normals | Lutein dipalmitate |
| Andreani & Volpi, 1956 | Normals (10) | Lutein dipalmitate |
| Mosci, 1956a | Normals | Lutein dipalmitate |
| Hayano, 1959a | Normals | Lutein dipalmitate |
| Wenzel et al., 2006 | Normals: No supplement (6); supplement (4) | 30 mg lutein + 2.7 mg zeaxanthin (12 weeks) |
| Rodriguez-Carmona et al., 2006 | Normal trichromats (24) | 10 mg (6 months) + 20 mg (6 months) of lutein or zeaxanthin, 10 mg lutein + 10 mg zeaxanthin or placebo |
| Kvansakul et al., 2006 | Normals (34) | 10 mg lutein, 10 mg zeaxanthin, 10 + 10 mg combination or placebo (6 months) |
| Bartlett & Eperjesi, 2008 | Normals (46) | 6 mg lutein + vitamins A, C, E + zinc + copper |
| Stringham & Hammond, 2008 | Normals (40) | 10 mg lutein + 2 mg zeaxanthin (6 months) |
MPOD indicates macular pigment optical density.
Abnormal light sensitivity, difficulty associated with glare, loss of contrast and slow dark adaptation are symptoms commonly reported by patients with hereditary retinal degenerations. It is possible that such symptoms could be attributable, at least in part, to the failure of MP to absorb scattered light, resulting in reduced contrast and definition along with excessive photoreceptor pigment bleaching by short wavelength light components.
The antioxidant and absorptive properties of MP would therefore suggest a potentially useful role for the macular carotenoids in retinal degenerations, where the clinical aim includes optimisation of current visual status in the short term and preservation of macular vision in the long term. Indeed, it is noteworthy that there have been reports (some dating back > 50 years) suggesting that patients with retinitis pigmentosa (RP) demonstrated improvements in visual performance following supplementation with lutein-containing compounds (reviewed elsewhere 16).
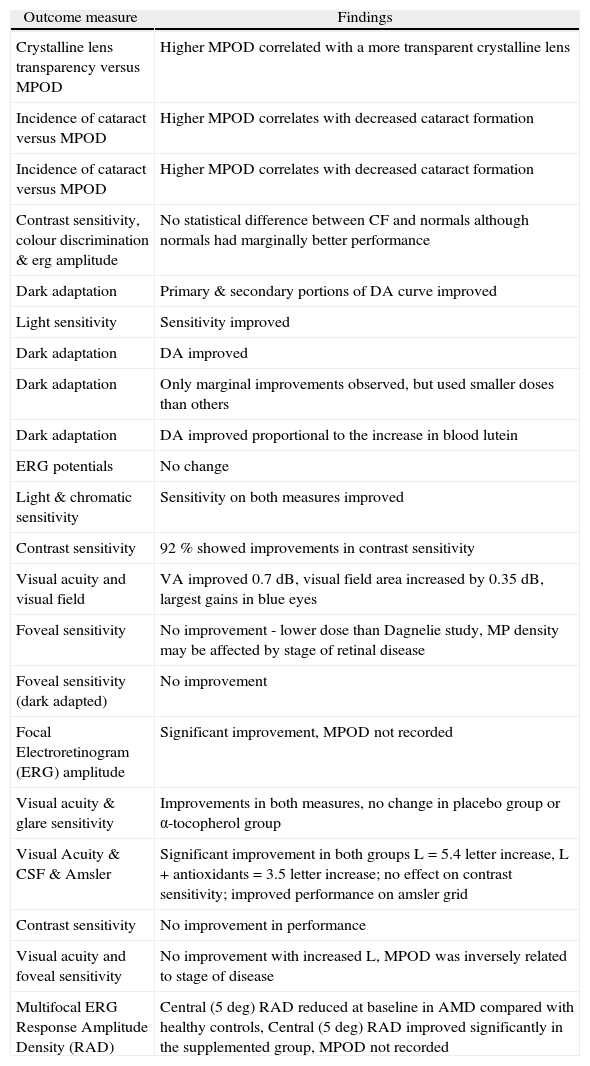
Dagnelie et al. 32 assessed the effect of L supplementation in patients with RP, and reported moderate visual improvements following short-term supplementation with L. Mean visual acuity improved by 0.7 dB and mean visual-field area by 0.35 dB, although the largest gains
| Outcome measure | Findings |
| Crystalline lens transparency versus MPOD | Higher MPOD correlated with a more transparent crystalline lens |
| Incidence of cataract versus MPOD | Higher MPOD correlates with decreased cataract formation |
| Incidence of cataract versus MPOD | Higher MPOD correlates with decreased cataract formation |
| Contrast sensitivity, colour discrimination & erg amplitude | No statistical difference between CF and normals although normals had marginally better performance |
| Dark adaptation | Primary & secondary portions of DA curve improved |
| Light sensitivity | Sensitivity improved |
| Dark adaptation | DA improved |
| Dark adaptation | Only marginal improvements observed, but used smaller doses than others |
| Dark adaptation | DA improved proportional to the increase in blood lutein |
| ERG potentials | No change |
| Light & chromatic sensitivity | Sensitivity on both measures improved |
| Contrast sensitivity | 92 % showed improvements in contrast sensitivity |
| Visual acuity and visual field | VA improved 0.7 dB, visual field area increased by 0.35 dB, largest gains in blue eyes |
| Foveal sensitivity | No improvement - lower dose than Dagnelie study, MP density may be affected by stage of retinal disease |
| Foveal sensitivity (dark adapted) | No improvement |
| Focal Electroretinogram (ERG) amplitude | Significant improvement, MPOD not recorded |
| Visual acuity & glare sensitivity | Improvements in both measures, no change in placebo group or α-tocopherol group |
| Visual Acuity & CSF & Amsler | Significant improvement in both groups L = 5.4 letter increase, L + antioxidants = 3.5 letter increase; no effect on contrast sensitivity; improved performance on amsler grid |
| Contrast sensitivity | No improvement in performance |
| Visual acuity and foveal sensitivity | No improvement with increased L, MPOD was inversely related to stage of disease |
| Multifocal ERG Response Amplitude Density (RAD) | Central (5 deg) RAD reduced at baseline in AMD compared with healthy controls, Central (5 deg) RAD improved significantly in the supplemented group, MPOD not recorded |
Duncan et al. 34 analysed MP levels and macular function in choroideremia (a progressive degeneration of photoreceptors, RPE and choroid). Once again, and in spite of augmented MPOD following supplementation, no improvement in retinal sensitivity was observed.
Aleman et al. 35 measured MPOD in patients with Stargardt's disease or cone-rod dystrophy with known or suspected disease-causing mutations in the ABCA4 gene, and investigated response to supplemental L in terms of changes in MPOD and central visual function. They reported that MPOD is inversely related to the stage of ABCA4 disease at baseline, and could be augmented by supplemental L in about two thirds of patients. However, measures of visual function, including visual acuity and foveal sensitivity, exhibited no discernable improvement after 6 months of supplementation. They concluded that the long-term influences of L supplementation on the natural history of such macular degenerations require further study.
Age-related macular degenerationAMD, as the leading cause of blindness in the western world, is the most commonly investigated retinal condition with respect to the potential benefits of supplemental L, Z, or meso-Z. Observations, including relative preservation of short wave sensitive cones centrally when compared to the perifoveal region 36 and the initiation of geographic atrophy in the perifovea, where MPOD is lowest, are consistent with the view that MP protects against AMD and against psychophysical changes known to precede this condition. Since publication of the findings of the Eye Disease Case-Control Study Group, where a 60 % risk reduction for AMD in association with a high dietary intake of L and Z was reported, 37 numerous investigators have further explored the relationship between dietary and serum levels of MP's constituent carotenoids and risk for AMD. 38 With a couple of exceptions (outlined below), studies investigating serum levels of, dietary intake of, or supplementation with, L and/or Z with respect to risk for AMD and/or its progression have (understandably) considered preservation, rather than enhancement, of visual performance, to represent the most appropriate outcome measure (reviewed elsewhere 39).
Richer 40 evaluated the effect of dietary modification on visual performance for patients with atrophic AMD. Fourteen male patients (70±9 years), receiving 0.73±0.45 portions of dark-green, leafy vegetables/day base intake, were placed on an additional portion of 5 ounces sautéed spinach 4 to 7 times per week or lutein-based antioxidant (3 subjects). Patients demonstrated short-term enhancement of visual function in one or both eyes in terms of amsler grid testing, Snellen acuity, contrast sensitivity, glare recovery, and subjectively on the Activities of Daily Vision Subscale. The authors concluded that the effect of dietary modification on the natural course of atrophic AMD warranted investigation in the context of a randomised, controlled trial.
Such an evaluation was conducted in the LAST (Lutein Antioxidant Supplementation Trial) study. Richer et al. 41 evaluated the effect of supplementation on visual performance in atrophic AMD on 90 subjects in a double blind, placebo controlled trial. Average MPOD increased by 0.09 log units (or 50 %) after 12 months, in the L and L plus antioxidant groups. The investigators observed concurrent and statistically significant improvements in contrast sensitivity, visual acuity and subjective measures of glare recovery in both treatment groups, but not in the control group. Snellen-equivalent acuity improved by 5.4 letters in patients supplemented with L, and by 3.5 letters in patients supplemented with L plus antioxidants, whereas improvements in contrast sensitivity were significantly better in the L plus antioxidant group than in the L group.
Falsini et al. 42 studied the effect of supplemental L on central retinal function, assessed electrophysiologically, in patients with early AMD, and reported a significant
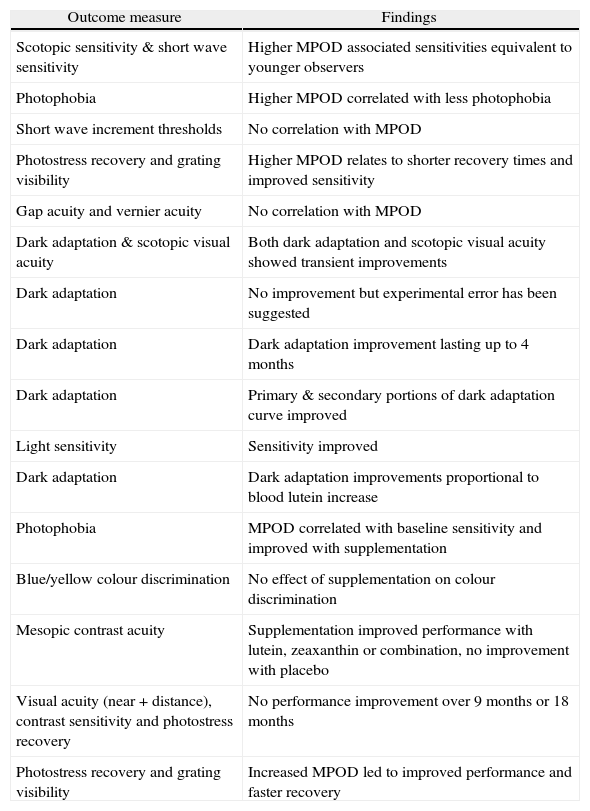
| Outcome measure | Findings |
| Scotopic sensitivity & short wave sensitivity | Higher MPOD associated sensitivities equivalent to younger observers |
| Photophobia | Higher MPOD correlated with less photophobia |
| Short wave increment thresholds | No correlation with MPOD |
| Photostress recovery and grating visibility | Higher MPOD relates to shorter recovery times and improved sensitivity |
| Gap acuity and vernier acuity | No correlation with MPOD |
| Dark adaptation & scotopic visual acuity | Both dark adaptation and scotopic visual acuity showed transient improvements |
| Dark adaptation | No improvement but experimental error has been suggested |
| Dark adaptation | Dark adaptation improvement lasting up to 4 months |
| Dark adaptation | Primary & secondary portions of dark adaptation curve improved |
| Light sensitivity | Sensitivity improved |
| Dark adaptation | Dark adaptation improvements proportional to blood lutein increase |
| Photophobia | MPOD correlated with baseline sensitivity and improved with supplementation |
| Blue/yellow colour discrimination | No effect of supplementation on colour discrimination |
| Mesopic contrast acuity | Supplementation improved performance with lutein, zeaxanthin or combination, no improvement with placebo |
| Visual acuity (near + distance), contrast sensitivity and photostress recovery | No performance improvement over 9 months or 18 months |
| Photostress recovery and grating visibility | Increased MPOD led to improved performance and faster recovery |
Bartlett and Eperjesi 43 undertook a prospective, 9-month, double-masked randomised controlled trial of the effect of supplementation with lutein combined with vitamins and minerals on contrast sensitivity among participants with age related maculopathy and atrophic AMD. Contrast sensitivity was assessed using a Pelli-Robson chart and participants were randomised into active and placebo treatment groups. The authors report no significant improvement in contrast sensitivity among either group and suggest that supplementation with 6mg/L and other antioxidant vitamins and minerals has no tangible benefit to this group (although one could argue that preservation rather than enhancement of performance might be a more suitable outcome measure for AMD patients) and further, that determination of optimum dosage levels requires further work. Their findings are naturally confined to the somewhat limited measure of contrast sensitivity with a Pelli-Robson chart that may not be best equipped to detect subtle changes in performance. Failure to record MPOD at baseline, and the low dosage of supplemental L, represent design flaws in that study, and limit the scope for meaningful comment. Parisi et al. 44 have also recently explored the influence of short-term carotenoid and antioxidant supplementation on electrophysiologically assessed retinal function in early AMD. Of the 27 early AMD patients enrolled in their study, 15 had daily oral supplementation of vitamin C (180mg), vitamin E (30mg), zinc (22.5mg), copper (1mg), lutein (10mg), zeaxanthin (1mg), and astaxanthin (4mg) for 12 months, while the remaining 12 had no dietary supplementation during the same period. Fifteen age-similar healthy controls were also assessed at baseline and followed-up for the duration of the study period without supplementation. Multifocal electroretinograms, in response to 61 M-stimuli presented to the central 20 degrees of the visual field (averaged across 5 retinal eccentricity areas between the fovea and mid-periphery: 0 degrees to 2.5 degrees, 2.5 degrees to 5 degrees, 5 degrees to 10 degrees, 10 degrees to 15 degrees, and 15 degrees to 20 degrees) were assessed at baseline in controls and in early AMD patients, and again at 6 months and 12 months. At baseline, they observed highly significant reductions of N1-P1 response amplitude densities (RADs) for the central five degrees surrounding the fovea in AMD patients when compared with healthy controls. For more peripheral retinal eccentricities, RADs were not significantly different from controls. After 6 and 12 months of treatment, the treated group showed highly significant increases in N1-P1 RADs for the two most central retinal areas, but not for more peripheral eccentricities beyond 5 degrees. The non-treated control group exhibited no significant RAD changes at any eccentricity. These findings suggest that in early AMD eyes, central retinal function (0 degrees -5 degrees) can be improved by supplementation with carotenoids and co-antioxidants. The study design, however, does not clarify whether such improvements in retinal function have a measurable impact on visual performance and experience, and the failure to measure and record MPOD somewhat limits the interpretation of these potentially important findings.
CataractOlmedilla et al. 45 investigated whether supplemental L influences visual function in patients with age-related cataract, where visual performance was evaluated by measures of visual acuity and glare sensitivity. This randomised, placebo-controlled trial revealed significant improvements in visual acuity and glare sensitivity following supplemental L, and the observed improvements were related to changes in serum levels of L, whereas no such improvements were observed in patients supplemented with placebo or with α-tocopherol. While contrast sensitivity was not recorded at baseline or during the supplementation phase, it is interesting to note that in cataract patients supplemented with lutein, contrast sensitivity at the end of the supplementation period was similar to or even better than that expected for control subjects of a similar age. The authors postulated that the observed improvements in the outcome measures were not the result of any change in the crystalline lens, but more likely to be the result of improved retinal function.
Studies in normal populationsPhotophobia and glarePhotophobia is a phenomenon experienced by all persons when illumination is suddenly and dramatically changed from dark to light, and is typified by the experience of switching on a bedroom light at night time. However, under normal daylight conditions, the experience of photophobia is somewhat more variable. Numerous clinical conditions (e.g. RP & AMD) are associated with photophobia, and, even in the absence of detectable disease, clinicians are often presented with patients whose primary complaint is of periodic or persistent sensitivity to bright light (but at levels which do not similarly affect colleagues/friends/family). Given its absorption characteristics, the optical density of MP may be important in determining an individual's threshold for the subjective complaint of photophobia.
Stringham et al. 46 explored the effect of the spectral composition of a target on visual discomfort, using electromyography and a rating scale to determine photophobia thresholds. They showed that, while there was a positive relationship between wavelength and the energy needed to produce photophobia for wavelengths between 520 and 640nm, at shorter wavelengths there was a notch centred at 460nm, the trough and shape of which resembled the log transmittance spectrum of MP. Their findings led the authors to suggest that MP may attenuate photophobia or visual discomfort induced by short wavelength sources.
These observations prompted a subsequent study investigating the relationship, if any, between MP and photophobia. 47 This two-part experiment explored the relationship between baseline MPOD levels and photophobia thresholds, as well as the effect of augmenting MPOD on such thresholds. Four subjects were supplemented with 30mg/L and 2.7mg Z daily for 12 weeks. Peak MPOD was observed to increase from 0.452 (± 0.11) at baseline to 0.536 (± 0.11) at the end of the period of supplementation. A significant and inverse relationship between baseline MPOD and threshold for photophobia was observed, such that individuals with higher MPOD had higher tolerance for short wavelength light. Furthermore, increasing MPOD over a 12-week period appeared to increase the threshold for photophobia for all subjects for short wavelength sources.
Recently, Stringham & Hammond have explored the influence of glare on visual performance, and how MPOD might influence any observed relationships. They first looked at baseline visual performance under glare conditions by evaluating photostress recovery (a sensitive indicator of macular function) and grating visibility. 48 The effect of veiling glare on grating visibility was explored using a five cycles per degree contrast grating stimulus, surrounded by a concentric annulus of adjustable intensity. For the photostress recovery test, the same stimulus was viewed following photostress with a 5 degree xenon white disc providing 5.5 log Trolands of retinal illuminance over 5 seconds’ duration. MPOD was a significant determinant of the deleterious effects of glare, with visual thresholds and photostress recovery times significantly and inversely related to MPOD. Further, high MPOD was associated with better visual performance in a way that was consistent with its spectral absorbance and spatial profile.
These observations prompted the same investigators to design and execute a trial of supplemental L (10mg per day) and Z (2mg per day), using the same testing conditions, but on this occasion looking for changes in performance associated with augmentation of MPOD. In this instance, they found that, following six months of supplementation, and an average increase in MPOD from 0.41 to 0.57, most subjects exhibited improved photostress recovery and glare tolerance in association with an increase in MPOD. More specifically, a 39 % increase in MPOD enhanced tolerance of intense glaring light by up to 58 % and reduced photostress recovery times by 14 %. 49
Although the authors wisely suggest a cautionary approach to the interpretation of their data and the wider implications of such findings, their conclusion that the results are “both large enough and sufficiently general to be meaningful in real life”, and that “supplementing L and Z could indeed be palliative for those suffering the consequences of glare”, is important and warrants further investigation in the form of a randomised clinical trial.
Spatial visionEngles et al. 50 have investigated the “acuity hypothesis”, exploring the relationship between MPOD and gap acuity and vernier acuity under “photopic” conditions. They report that neither gap acuity nor vernier acuity were significantly related with MPOD, and concluded that their “data suggest that the predictions of the acuity hypothesis do not hold”. While the authors qualify their findings as appropriate to their specific testing conditions alone, several study limitations (other than those recognised by the authors) warrant brief discussion.
Firstly, although the authors report that their conclusions are relevant for photopic conditions, their adopted background luminance levels are in the low photopic range at best (17cd/m2 for the achromatic condition, and 15.7 cd/m2 for the chromatic condition), and certainly not appropriate for evaluation of photopic visual function. Indeed, given the subtle nature of any performance changes likely to be facilitated by MP, the background luminance difference (≈8 %) between the two testing conditions is also a potentially confounding factor.
Secondly, while all subjects were corrected to 6/6, it is plausible, indeed probable, that the actual acuity limits of their study population ranged widely between the 6/6 level employed up to a likely 6/3 limit for a young healthy subject. This potential two-fold range in acuity, subserved by individual optical, anatomic and neural architectures, would have a strong influence on both gap and vernier acuity tasks, almost certainly more powerful than MPOD. Also, by adopting a 6/6 limit, the investigators most likely failed to correct for potentially significant amounts of uncorrected axial astigmatism in some subjects, which could significantly influence performance on both of the chosen tasks (testing of vernier and gap acuity limits). While the authors could argue that any such variables remained consistent between testing conditions, we believe it would be more appropriate to eliminate sources of variability such as residual refractive error, so that all subjects operate at their limits of acuity.
The adoption of a single spatial frequency and contrast setting further limits the conclusions that can be drawn from this paper. The effect of MP, for example, might differ significantly under different spatial frequency and or contrast ranges. Assessment of visual performance across the full contrast sensitivity function might represent a more thorough and rigorous assessment of MP's capacity to affect visual performance through attenuation of the effects of chromatic aberration and light scatter.
Finally, the subjects employed in the Engles study typically exhibited average to high MP levels, with few subjects exhibiting MP levels below 0.20. Reading and Weale 15 previously modelled the potential effect on MP in terms of attenuation of the effects of chromatic aberration, and suggested that, due to the non-linear nature of the effect, MPOD levels above 0.30 were probably superfluous. Based on the assumptions of this model, a study on the effect of MP on visual performance might require the inclusion of relatively more subjects that exhibit low MPOD levels inorder to demonstrate an effect.
These limitations of the cited study serve to emphasize the challenges inherent in investigating the role of MP in visual performance and experience, which rest on the need (insofar as is possible) to disentangle the influence of MP from the often unquantifiable and variable influences of individual optical and neural architectures.
Loughman et al., 51 in a cross sectional analysis involving some 142 young healthy subjects, observed statistically significant relationships between MPOD and best corrected visual acuity, and also with photopic and mesopic contrast sensitivity at intermediate spatial frequencies. MP appeared to contribute to up to a 0.1 log unit refinement of high contrast visual acuity (equivalent to one extra line on the acuity chart, or the effective correction of up to 0.25D or residual blur). The correlations between MPOD and visual acuity and contrast sensitivity however, although statistically significant, account for only a small percentage of the potential variability (r2 values < 10 %), so should be interpreted cautiously with respect to its clinical relevance in the absence of a more rigorous placebo controlled supplementation study.
Bartlett and Eperjesi 52 set out to explore the effect of L supplementation on visual performance among healthy observers. Similar to their AMD trial (2007), the authors report no effect of supplementation on performance measures ranging from distance and near visual acuity, contrast sensitivity and photostress recovery. The results are somewhat unsurprising however given (a) the low dose, 6mg/L supplement used, (b) the basic nature of the series of tests employed to evaluate visual performance, and (c) the small number of subjects tracked over 9 months (n = 46) and 18 months (n = 29) across such a broad age range (22-73 years). Once again, their failure to record MPOD or serum L and Z levels means that only qualified comment can be made as to the significance of the reported findings.
Armstrong et al., in a primitively designed pilot study (involving only one subject) presented at a recent conference (ARVO 2008, Poster # 4964/D984), evaluated macular function on a serial basis throughout a 4-month period of supplementation with L and Z. Looking at a series of psychophysical and electrophysiological outcome measures, they evaluated the effect of supplementation on dark-adapted thresholds and recovery kinetics, pattern visual evoked potentials (PVEPs) [before and after photostress], and PERG amplitude. An MPOD increase of approximately 33 % was accompanied by a 23 % improvement in 650nm dark adapted thresholds (from 30dB to 37dB) and by an increase in PERG amplitude, but not by a change in cone recovery kinetics or photostress PVEP recovery. Although these findings should be interpreted with caution, particularly given that only one subject was tested, they are again suggestive of an improvement in macular function following augmentation of MPOD in young healthy subjects.
The inconsistencies in spatial vision data with respect to MPOD reflect the difficulty inherent in isolating performance tasks which may be influenced by MP. Furthermore, the wide inter-individual variability of MPOD 53 renders the interpretation of such studies all the more challenging, particularly where such investigations depend on cross-sectional rather than longitudinal data. It would however seem to be the case that, as far as spatial vision is concerned, the effect, if any, of MP on performance appears small, at least for individuals with average to high MPOD.
Colour visionSince the MP absorption spectrum ranges from about 400 to 520nm and peaks at 460nm, 54 it would seem likely that these pigments influence colour vision through selective absorption of short wavelengths, thereby influencing the short-wave sensitive (SWS) cones and the blue-yellow opponent-colour channel. Moreland and Dain 55 (1995) reported that hue discrimination, measured using the Farnsworth-Munsell 100-Hue test (FM100), is indeed adversely affected (primarily) for blue wavelengths, by simulation of high MPOD using liquid notch filters containing carotene in a benzene solution. Comparing the results with those obtained with a neutral filter, they concluded that this effect was not simply the result of reduced retinal illuminance. Further evidence supporting an effect of MPOD on short wavelength vision has been obtained from studies of SWS cone sensitivity. 56,57 It has also been shown that colour discrimination measured by a colour matching technique is influenced by MPOD. 58,59
However, two recent studies using alternative methods, produced conclusions differing from those of the above mentioned studies. Firstly, a study of the effects of dietary supplementation with macular carotenoids on MP found no correlation between the level of MP (measured by heterochromatic flicker photometry) and red-green (RG) or yellow-blue (YB) colour discrimination thresholds, though it was reported that RG vision was improved following supplementation. 60 Secondly, RG cancellation profiles have been reported to be highly correlated with MPOD, while profiles for YB were independent of both eccentricity and MPOD. 61 Further support comes from a study of anomaloscope Moreland match midpoint data, in which no difference was reported between post-cataract patients with blue-absorbing intra-ocular lenses (IOLs) and those with clear IOLs. 62
Thus, the influence, if any, of MP on colour vision remains uncertain at the present time. However, It is possible that an artificial filter creates short-term changes in colour vision and that an autoregulatory process adjusts retinal and/or cortical colour mechanisms on a long-term basis in response to an individual's naturally occurring MPOD. 61 This hypothesis is supported by data showing a consistent shift in achromatic locus over a three month period for post-surgical cataract patients, 63 and by evidence of plasticity of adult neural colour mechanisms. 64
Preservation of ‘youthful’ vision into old ageIn the elderly, pre-retinal image degradation and slower encoding results in featurally-compromised representation of spatially-extended search arrays. Even with appropriate optical correction, older adults therefore do not possess the spatial resolving power of the young adult. Such losses are not confined to high spatial frequencies, but contrast sensitivity losses are observed across a range of intermediate frequencies. 65 Indeed, many changes in both structure and function of the visual system, such as pupillary miosis and loss of crystalline lens transparency, 66 accompany the aging process (summarised elsewhere 67). The consequence of such changes is a reduction in retinal illuminance, such that equiluminant stimuli do not result in equal retinal illuminance for different age groups. Human visual performance therefore tends to decrease with age (Figure 4). Such effects are to some extent unavoidable, and a natural consequence of aging.

Effect of normal aging on contrast sensitivity. Experimental data show a 1-log unit sensitivity decrease from age 60 to 95.
The most significant role of MP in vision, however, may rest on the potential of L, Z and meso-Z to retard the aging process through their antioxidant properties. It is important to note that MP acts, uniquely, as an antioxidant, both passively and actively, the former mechanisms being dependent on its ability to limit photo-oxidative damage by filtering short wavelength light at a pre-receptorial level and the latter mechanism attributable to its capacity to quench reactive oxygen intermediates.
The inter-individual variability in MPOD, consistently observed in cross-sectional studies, may have important implications for the long term health and viability of the central retina. In subjects with little MP, the cumulative and chronic effects of increased exposure of photoreceptors to short wavelength light, coupled with a weaker local capacity to quench free radicals, could, in theory at least, accelerate the onset of physiological and pathological aging of the retina.
In support of such a notion, Hammond et al. 56 have shown that high MPOD was associated with the retention of youthful scotopic and short wave sensitivity and suggested that MP may retard an age-related visual decline. The potential benefits of increased MPOD appear not to be confined to the retina. Hammond et al. 68 reported a positive and significant association between crystalline lens transparency and MPOD, and speculated that high concentrations of the macular carotenoids in the lens probably accompany high concentrations at the macula, and protect against the effects of oxidation in the lens (thereby maintaining transparency). Indeed, other studies have shown an association between a high dietary intake of L and Z with decreased incidence of cataract formation. 69,70
Werner and Steele 71 demonstrated age-related sensitivity losses of foveal colour mechanisms across all three cone types, although the sensitivity loss for short wavelength sensitive cones (S-cone) was lower (at 0.08 log units per decade), when compared to 0.11 log units loss per decade for both medium (M-cone) and long wave (L-cone) cones. Werner et al. 57 later explored the senescence of foveal and parafoveal cone sensitivities and their relation to MPOD. Again, they report age-related decline of foveal and parafoveal increment thresholds. Interestingly however, and consistent with the hypothesis that the MP protects the photoreceptors from senescent losses in sensitivity, a significant and positive correlation was found between foveal MPOD and differential S-cone log sensitivity losses at the fovea and at the parafovea, but not with differential M- and L- cone log sensitivity losses at the retinal loci. This finding, however, was independent of age, prompting the authors to postulate that it was due to local gain changes, resulting from differential filtering of incident light by the MP between the fovea and the parafovea.
Haegerstrom-Portnoy 72 also examined S-cone versus L-cone sensitivity in a group of young and older adults to determine whether MP protects the human fovea from retinal neural damage caused by visible-light exposure over a lifetime. While there was no difference observed for L-cone sensitivity between groups, the older group showed a significant differential loss of S-cone sensitivity across the retina compared with the younger group, with greater loss of sensitivity at non-foveal locations than at the fovea. This observation is again suggestive of a protective effect of MP on foveal function.
Schupp et al. 73 endeavored to explore the hypothesis from a different perspective, postulating that if high levels of MP might forestall the effects of normal aging, then low levels of MP might accelerate the normal aging process. Cystic fibrosis (CF) is a condition associated with defective gastrointestinal absorption of carotenoids as a result of pancreatic insufficiency. Low serum concentrations of carotenoids, including the constituents of MP, are invariably reported in CF patients. Given the repeatedly observed positive and significant relationship between MPOD and serum concentrations of its constituent carotenoids (reviewed elsewhere 74), it can be reasoned that patients with CF would have low MPOD. Schupp et al. 73 assessed visual performance in ten cystic fibrosis patients, in whom serum concentrations of L and Z and MPOD were predictably and significantly lower than control subjects, and typically less than 50 % of the values observed amongst control subjects. However, visual performance (contrast sensitivity, colour discrimination and multifocal ERG amplitudes) were statistically similar for CF patients and control subjects.
While the basic rationale of this study is provocative, there are however a number of concerns with the methodology. With six of the ten CF subjects aged between 21-27 years, it is unlikely that such a youthful population sample would demonstrate accelerated aging effects on visual function (even in the presence of chronically low MPOD levels). In any case, given the theoretical possibility that higher levels of MP might be associated with enhanced visual performance, it is unclear from this publication as to how functional differences, which might have been observed between the CF and control groups, could be attributable to age effects rather than simply to differences in MPOD. The authors conceded that a longitudinal assessment of an older CF population is required to address the hypothesis more appropriately.
Hammond and Wooten 75 investigated the relationship between MP, critical flicker fusion frequency (CFF) and age, citing CFF as a general measure of visual health. They found a significant decline in CFF values with age. There was a significant and positive relationship however between MPOD and CFF values that was independent of age. The authors conclude that these results are consistent with a protective effect of MP on visual health across the lifespan. While such investigations appear to be at a very early stage, preliminary results suggest a role for MP in temporal vision and, specifically, that high MPOD may protect the retina and defer some typical age-related changes in temporal vision.
ConclusionsVisual performance in the normal human is less than ideal, and it has been shown that visual performance improves once chromatic and monochromatic aberrations are removed. 76 As a consequence, numerous interventions which attenuate these aberrations have been developed in an attempt to optimise and/or enhance visual performance, Wavefront-guided laser refractive eye surgery, wavefront-guided spectacle lenses, short wavelength-filtering intraocular lens implants, short wavelength-filtering contact lenses and short wavelength filtering spectacle lenses are all directed towards improving or optimising visual performance. These techniques, however, are primarily intended for persons with pre-existing ocular abnormality or disease, and there has been a conspicuous lack of concerted effort to improve (or maintain) visual performance in subjects without demonstrable ocular patho logy. Augmentation of MPOD by means of supplementation remains a plausible and realistic means (in theory at least) of optimising and/or enhancing visual performance in a normal population.
Future studies should address the issue of whether variations in MPOD relate to visual performance, and whether high MP levels can preserve or prolong optimal central visual function into old age. Indeed, some studies have reported that high levels of MP are associated with preservation of retinal sensitivity in the elderly.
MP has ideal properties, in terms of location and spectral absorbance, to be beneficial for visual performance and experience. Longer life expectancy, increased exposure to short wavelength light (ancestors had little or no short wavelength light exposure after dark), increased effects of scatter from expanding smog and haze, modern visual requirements and the ever-increasing incidence of AMD heightens the importance of both optimising (and possibly enhancing) visual performance in the working population, and preserving such performance into old age. Robust evidence, in support of the psychophysically plausible rationale, that MP contributes to visual performance and experience in a favourable way is, however, still lacking. The findings of the studies cited above, whether demonstrating a benefit of MP to visual performance and experience or not, should be interpreted with full appreciation of their design limitations, and it should be understood that a cross-sectional study represents an inappropriate design to investigate fully any contribution that MP makes to visual performance. It is unwise to assume that the role of MP in visual performance, if any, can be easily studied, given the multitude of typically individual and occasionally enigmatic factors that influence our visual experience.
Given the numerous optical and neural factors that influence and dictate visual performance, and the consequential and associated difficulties in isolating improvements in visual performance, any study designed to investigate the influence of MP in this regard should include questionnaire-based analyses of subject perceptions of personal visual experience. Such an approach will facilitate investigation of the potential role of MP in visual performance in the real world, in a natural and ever-changing environment, which is often poorly reflected in our current and limited arsenal of testing modalities. None of the studies which reported a beneficial effect of MP augmentation adequately address the question of (1) whether such increases in MPOD and the observed psychophysical functional improvements translate into tangible improvements in visual experience outside the laboratory or (2) whether such improvements can be longitudinally maintained to preserve functional perfor mance and experience into old age.
Because of the inter-individual variability in MPOD and psychophysical visual function, a study designed to investigate the contribution of MP to visual performance and experience should be able to study the relationship between changes in these parameters within subjects over time, and only a study where MP is augmented by supplementation and/or dietary modification can meet this essential criterion. Interestingly, of the studies cited in this review, there appears to be one reasonably consistent finding, despite varied design limitations, studies involving supplementation among normal and diseased eyes typically report measurable benefits in terms of visual performance, in terms of photophobia thresholds, glare sensitivity, dark adapted thresholds, PERG amplitudes and mesopic contrast sensitivity among others.
Thus far, there appears to be little or no evidence of any adverse effect of higher levels of MP on visual performance. In a study designed to determine the influence of macular pigment absorption on blue-on-yellow perimetry, Wild and Hudson 77 found that the net effect of ocular media and MP absorption relative to 460nm was to attenuate the blue-on-yellow visual field at the fovea by approximately 0.80 log units and elsewhere by 0.40 log units, the difference being attributable to MP. Unpublished results from our own laboratory suggest no association between MPOD and colour matching or colour discrimination ability, although we have observed a non-significant inverse association between central short wavelength sensitivity and MPOD (data on file). The possibility of an adverse effect of MP augmentation on colour vision, short wavelength sensitivity and other functional measures does merit future investigation.
The optical, physiological and neurological interactions that contribute to vision suggest that the optimal level of MPOD, from a performance perspective, may be personal to an individual eye. In other words, and for example, even if MP is found to be important for visual performance and experience, exceeding a particular optical density of the pigment may yield no further measurable or appreciable advantage, and this level may vary substantially from one individual to the next. It is also important to note that testing conditions are often incapable of reflecting more natural environments, and any observed absence or presence of MP's contribution to visual performance and experience may not necessarily hold true in a natural environment (for example, against the background of a bright blue sky).
Although it remains difficult to draw firm conclusions regarding the relationship between MP and visual performance, certain patterns do appear to exist. In normal observers, the effect on spatial and colour vision appears small in comparison to the observed effects on photophobia and glare sensitivity, while, in subjects with established eye disease, there appears a relatively consistent beneficial effect of MP supplementation on visual performance. Any effects observed, whether through optical or biological mechanisms, may also be magnified when increased emphasis is afforded to those with chronically low MPOD levels. We need and should support an appropriately powered, randomised, controlled trial, which is designed to further evaluate whether visual performance and experience can be optimised or enhanced, or indeed adversely affected, with supplemental macular carotenoids.
Conflict of interestThe authors state they have no conflict of interest.
The authors would like to thank Larry N. Thibos for assistance in the production of Figure 2.





